

What is caffeine?
Caffeine is a psychoactive drug that exerts a powerful stimulant effect on the central nervous system. It is naturally found in coffee, tea, and chocolate, but is also added to some soft drinks (e.g. colas), energy drinks and energy shots.[8] The National Health and Nutrition Examination Survey (NHANES) found that roughly 89% of adults in the US consume caffeine daily.[12] People often use caffeine-containing beverages to help restore alertness and wakefulness or to reduce drowsiness when feeling mentally fatigued.[13][14] Caffeine is also available as a purified powder, and it is found in some dietary supplements claiming to cause weight loss or increase energy (e.g. pre-workouts).[8][15][16] Additionally, caffeine is found in some over-the-counter drugs used to relieve pain[17][18] and in prescription drugs used to treat breathing problems in premature babies.[19]
After ingestion, caffeine acts rapidly on the central nervous system, triggering several short-lived effects including an increase in heart rate, blood pressure, and urine output.[20][21][22][23] However, with regular daily use, these symptoms of caffeine typically become less pronounced.[20][22][8][13][14]
What are caffeine’s main benefits?
Caffeine restores alertness and wakefulness, and reduces drowsiness during mental fatigue.[24][25][26] It also improves several aspects of cognitive function — accuracy, reasoning, memory, reaction time, attention, etc. — during cognitively and/or physically demanding tasks.[27][24][25][26] Furthermore, caffeine can improve some aspects of cognitive function that have been impaired by acute sleep deprivation.[25][26]
Caffeine can also help with pain relief. For example, when caffeine is added to analgesic (pain relieving) drugs like acetaminophen (also known as paracetamol) or ibuprofen, there is a small but clinically meaningful improvement in pain.[17][18]
In hospitals, caffeine (typically caffeine citrate) is used to treat apnea (temporary cessation of breathing) in premature babies, but both the optimal dosing strategy and caffeine’s effect on survival and subsequent neurocognitive development are unclear.[19] Furthermore, in people with asthma, caffeine may have a small beneficial effect on some aspects of respiratory function (forced expiratory volume in one minute, FEV1, and mid-expiratory flow rate) for up to four hours.[28] However, this effect also has implications for asthma diagnostics because prior caffeine ingestion may reduce the accuracy of lung function tests.[28]
Caffeine can raise whole-body fat oxidation rates, both at rest and during exercise,[29][30] and increase daily energy expenditure,[31][32] but these effects are negligible — approximately 400 kilojoules or 100 kilocalories per day. The effect of caffeine on appetite regulation and energy intake is less clear and varies depending on factors like timing and dose.[32][33] Consequently, the role of caffeine in weight loss and/or weight management is unclear. Despite one meta-analysis concluding that caffeine can promote reductions in weight, BMI, and body fat,[34] its utility is limited because the included trials used energy-restriction diets and/or ephedrine, a powerful stimulant that causes weight loss.
Caffeine is widely used in sports because it can improve both aerobic and anaerobic performance in trained and untrained individuals.[1] For example, when taken before or during exercise, caffeine has small to moderate-sized effects of lowering the rating of perceived exertion (RPE)[35][36] and improving aerobic endurance, anaerobic power, sprint speed, muscle endurance, muscle strength, muscle power (jump height), and agility.[1][37][38] These effects are found across a range of sports (running, cycling, and swimming, etc.) including skill-based team sports (e.g. basketball, soccer, etc.).[1][39][40] Furthermore, when taken before and/or during exercise, caffeine may also improve cognitive functions, such as attention, reaction time, memory, and feelings of fatigue.[24] One drawback is the predominance of research studies that only included young male participants.[37][1] Some meta-analyses find similar performance benefits in females to those found in males,[41][42][43] but more randomized controlled trials in females are needed to substantiate the generalization of sports nutrition recommendations for caffeine.[1][44]
Do different genotypes influence the effect of caffeine?
Does regular caffeine use reduce the benefit of caffeine on exercise performance?
Does regular caffeine use reduce the risk of cognitive disorders like dementia and Alzheimer's disease?
Is caffeine a banned substance for athletes?
Does coffee have the same effect as isolated caffeine on exercise performance?
Does using a caffeine-containing mouth rinse or chewing a caffeine gum have the same effect on exercise performance as ingesting caffeine?
What are caffeine’s main drawbacks?
Caffeine is generally safe for most people if used within the recommended amounts (up to 400 mg per day in a healthy adult, or up to 200 mg in a single dose — see the dosage information section.[3] Furthermore, caffeine-containing beverages like tea and coffee are socially acceptable and widely used.[45][12] However, caffeine has several side effects and drawbacks.
The side effects of caffeine include a short-lived rise in heart rate and blood pressure, heart palpitations, headache, increased urine output, nervousness, gastrointestinal problems, etc.[20][21][22][23] Caffeine can also raise heart rate during and following exercise.[46][2] Some of these side effects, particularly those related to heart rate, blood pressure, and urine output, may subside with regular use due to increased tolerance.[20][22][8][13][14] Because caffeine can cause mild drug dependence, some people also experience withdrawal symptoms — headache, drowsiness, and irritability — when they stop using caffeine after regular daily intake.[47]
One of the major drawbacks is that caffeine impairs sleep quality and can reduce sleep duration.[48][49] A recent meta-analysis found that to prevent deleterious effects on subsequent sleep, coffee should be consumed at least 9 hours before bedtime, and a caffeine-containing pre-workout supplement should be consumed at least 13 hours before bedtime.[48]
Sadly, there have been several case reports of caffeine toxicity[50][51][52][53] and caffeine-related death.[54][55][56][57] However, considering the high global prevalence of caffeine intake, these cases are rare.[45][12][58] Such cases also typically involve exceptionally high caffeine intakes (usually unintentional but sometimes purposeful) or caffeine intake combined with other drugs and alcohol.[54] Some instances of toxicity and death involve energy drinks — beverages that contain as much as 300 mg of caffeine plus other stimulants like taurine, in addition to B vitamins and sugar.[59] At the population level, energy drinks are unlikely to cause harm because there is a low prevalence of consumption.[60][61] However, in certain groups (children, teenagers, people with underlying heart conditions), health risks can emerge when people far exceed recommended daily caffeine intake limits by consuming energy drinks with other caffeine-containing foods and beverages.[60][61]
Some epidemiological studies have found a relationship between high daily caffeine intake (e.g. more than 4 to 6 cups of coffee per day) and increased cardiovascular disease risk.[62][63][64][65] However, the evidence from mendelian randomization studies and meta-analyses of epidemiological studies shows that caffeine (or coffee) intake does not increase the risk of cardiovascular disease, including coronary artery disease, stroke, heart failure, and cardiovascular disease mortality,[61][65][66][67] and that moderate intake might even be protective.[68][69][70][71]
Some studies also find a relationship between caffeine intake during pregnancy and poorer health outcomes for mother and baby following birth; however, the entirety of the evidence is equivocal.[72][73] Furthermore, it is unclear whether reducing caffeine intake during pregnancy has any effect on pregnancy outcomes, including infant health and birth weight.[72][73] However, some authorities recommend that pregnant women limit their caffeine intake to 200 milligrams per day (the equivalent of 1–2 cups of coffee per day or less).[5]
How does caffeine work?
Caffeine’s mechanisms of action are not completely understood, but the proposed mechanisms likely act in unison.[14][1][74]
Caffeine’s primary action is in the central nervous system, where it prevents adenosine from binding to its receptor.[75][14][74] This affects the secretion of several neurotransmitters — norepinephrine, dopamine, acetylcholine, serotonin, GABA, etc. — involved in alertness, mood, motivation, memory, and pain perception.[14][76] For example, adenosine released during exercise decreases the secretion of dopamine, a neurotransmitter that increases mood and motivation.[1][77] So, when taken before or during exercise, caffeine allows dopamine secretion to continue, potentially maintaining mood and motivation to work hard.[77] This, coupled with lower pain perception caused by the lack of adenosine signaling, may partly explain caffeine’s sports-performance-enhancing effects.
Caffeine also plays a role in cellular calcium transport, which regulates neurotransmitter secretion in neurons and contractile force in muscle cells.[14][74] Caffeine can have a direct effect on calcium transport in muscle cells and may, therefore, have a direct effect on muscle contractile force,[1][78][79][80][81][82] providing an additional explanation for its performance-enhancing effects.
Caffeine can also inhibit a group of enzymes called cyclic nucleotide phosphodiesterases (PDEs), preventing the breakdown of cyclic adenosine monophosphate (cAMP).[14][74] This is important because cAMP stimulates the release of neurotransmitters like dopamine, epinephrine, and norepinephrine, which regulate aspects of cognitive function, including alertness, mood, motivation, and memory.[14][74] However, this mechanism requires more research and may only occur in the presence of high concentrations of caffeine, i.e., following high caffeine doses.
What are other names for Caffeine
- Coffee extract
- Tea extract
- 1 3 7-Trimethylxanthine
- Caffeic acid
Dosage information
For sports performance
The optimal dose for sports performance benefits is 3–6 mg per kilogram of bodyweight (approximately 200–400 mg in a 70 kg person), taken around 60 minutes before exercise.[1] Using a dose at the low end of this range, approximately 3 mg/kg of caffeine before exercise (approximately 200 mg in a 70 kg person), achieves the benefits along with the lowest risk of side effects.[2]
Safety in adults
Caffeine is generally safe for most people if used within the recommended amounts.[3][4] The US Food and Drug Administration (FDA) and the European Food Safety Authority (EFSA) report that a daily total caffeine intake of up to 400 milligrams (mg) does not raise safety concerns for non-pregnant adults.[5][6] The EFSA further reports that a daily total caffeine intake of up to 200 mg in pregnant and lactating women does not increase safety concerns for unborn babies or breastfed infants, and that a single caffeine dose of up to 200 mg (approximately 3 mg per kg bodyweight for a 70-kg adult) does not raise safety concerns.[5] In other words, caffeine intake is unlikely to cause detrimental health effects if daily intake is below 400 mg/day in non-pregnant adults, or below 200 mg/day in pregnant/lactating women, and if a single dose is less than 200 mg.
Safety in children and teens
In children and adolescents, more research is needed to clarify safety, health risks, and tolerable upper intake limits.[5] Evidence from a systematic review suggests that children and adolescents should consume no more than 2.5 mg per kg of bodyweight per day (equivalent to 100 mg for a 40 kg person).[3] But some organizations discourage all consumption of caffeine and other stimulants by children and adolescents.[6]
Caffeine in food and drink
Caffeine concentrations in various foods — coffee, tea, soft drinks (cola), chocolate, energy drinks, etc — are highly variable.[5] For example, a cup of coffee may contain approximately 50–200 mg of caffeine, but the amount per cup is highly variable depending on the type of bean, how it is roasted, and how it is prepared.[7][8] Some energy drinks contain very high amounts of caffeine, and the FDA recommends avoiding dietary supplements that contain pure or highly concentrated caffeine because a single teaspoon of pure powdered caffeine contains about 5 grams (5000 milligrams), far higher than the upper recommended limit of daily intake and approaching the toxic dose.[9]
For a detailed overview of caffeine dosing, read our article “How Much Caffeine Is Too Much?”.
Caffeine in caffeine citrate
Note that caffeine citrate is about 50% caffeine by weight, i.e., a 50 mg dose of caffeine citrate contains approximately 25 mg of caffeine.[10][11]
Examine Database: Caffeine
Research FeedRead all studies
In this crossover study, both an espresso mouth rinse and caffeine gum improved some metrics of aerobic capacity but not power production compared to a placebo.
Frequently asked questions
Caffeine is a psychoactive drug that exerts a powerful stimulant effect on the central nervous system. It is naturally found in coffee, tea, and chocolate, but is also added to some soft drinks (e.g. colas), energy drinks and energy shots.[8] The National Health and Nutrition Examination Survey (NHANES) found that roughly 89% of adults in the US consume caffeine daily.[12] People often use caffeine-containing beverages to help restore alertness and wakefulness or to reduce drowsiness when feeling mentally fatigued.[13][14] Caffeine is also available as a purified powder, and it is found in some dietary supplements claiming to cause weight loss or increase energy (e.g. pre-workouts).[8][15][16] Additionally, caffeine is found in some over-the-counter drugs used to relieve pain[17][18] and in prescription drugs used to treat breathing problems in premature babies.[19]
After ingestion, caffeine acts rapidly on the central nervous system, triggering several short-lived effects including an increase in heart rate, blood pressure, and urine output.[20][21][22][23] However, with regular daily use, these symptoms of caffeine typically become less pronounced.[20][22][8][13][14]
Caffeine restores alertness and wakefulness, and reduces drowsiness during mental fatigue.[24][25][26] It also improves several aspects of cognitive function — accuracy, reasoning, memory, reaction time, attention, etc. — during cognitively and/or physically demanding tasks.[27][24][25][26] Furthermore, caffeine can improve some aspects of cognitive function that have been impaired by acute sleep deprivation.[25][26]
Caffeine can also help with pain relief. For example, when caffeine is added to analgesic (pain relieving) drugs like acetaminophen (also known as paracetamol) or ibuprofen, there is a small but clinically meaningful improvement in pain.[17][18]
In hospitals, caffeine (typically caffeine citrate) is used to treat apnea (temporary cessation of breathing) in premature babies, but both the optimal dosing strategy and caffeine’s effect on survival and subsequent neurocognitive development are unclear.[19] Furthermore, in people with asthma, caffeine may have a small beneficial effect on some aspects of respiratory function (forced expiratory volume in one minute, FEV1, and mid-expiratory flow rate) for up to four hours.[28] However, this effect also has implications for asthma diagnostics because prior caffeine ingestion may reduce the accuracy of lung function tests.[28]
Caffeine can raise whole-body fat oxidation rates, both at rest and during exercise,[29][30] and increase daily energy expenditure,[31][32] but these effects are negligible — approximately 400 kilojoules or 100 kilocalories per day. The effect of caffeine on appetite regulation and energy intake is less clear and varies depending on factors like timing and dose.[32][33] Consequently, the role of caffeine in weight loss and/or weight management is unclear. Despite one meta-analysis concluding that caffeine can promote reductions in weight, BMI, and body fat,[34] its utility is limited because the included trials used energy-restriction diets and/or ephedrine, a powerful stimulant that causes weight loss.
Caffeine is widely used in sports because it can improve both aerobic and anaerobic performance in trained and untrained individuals.[1] For example, when taken before or during exercise, caffeine has small to moderate-sized effects of lowering the rating of perceived exertion (RPE)[35][36] and improving aerobic endurance, anaerobic power, sprint speed, muscle endurance, muscle strength, muscle power (jump height), and agility.[1][37][38] These effects are found across a range of sports (running, cycling, and swimming, etc.) including skill-based team sports (e.g. basketball, soccer, etc.).[1][39][40] Furthermore, when taken before and/or during exercise, caffeine may also improve cognitive functions, such as attention, reaction time, memory, and feelings of fatigue.[24] One drawback is the predominance of research studies that only included young male participants.[37][1] Some meta-analyses find similar performance benefits in females to those found in males,[41][42][43] but more randomized controlled trials in females are needed to substantiate the generalization of sports nutrition recommendations for caffeine.[1][44]
A genotype describes a variant of a gene that can alter the function of the protein the gene is translated into. There are specific proteins (enzymes) in the body that metabolize caffeine and there are several variants of the genes that produce those enzymes. One enzyme, cytochrome P450 1A2, which is responsible for approximately 95% of caffeine metabolism, is coded by the CYP1A2 gene. Different variants of this gene cause people to be “slow” or “fast” metabolizers of caffeine. The current evidence shows that variants in the CYP1A2 gene may influence habitual caffeine intake[83][84] and caffeine’s effect on cognitive function.[85] While some studies implicate CYP1A2 gene variants in the effect of caffeine on sports performance, the effects are highly variable and further high-quality studies are needed.[86][87]
Variants of the ADORA2A gene, which codes the adenosine A2A receptor protein, affect caffeine’s ability to prevent adenosine binding to adenosine receptors. This causes people to have “high” or “low” caffeine sensitivity. The current evidence shows that variants in the ADORA2A gene may influence the anxiogenic (anxiety-causing)[83][85] and sleep-disturbing[85][88][89][90] effects of caffeine.
Some studies find a relationship between different CYP1A2 and ADORA2A genotypes and cardiometabolic risk factors, like blood glucose responses to a meal and blood pressure.[91] However, the current evidence suggests that different genotypes do not influence the effect of caffeine on cardiovascular disease risk.[63][65]
Some studies show that sustained daily use of caffeine can blunt some of the physiological responses following caffeine intake,[92][20][22][8][13][14] and may reduce its exercise performance-enhancing effects.[93][94][95] Therefore, habitual caffeine users may experience a lower performance-enhancing effect than nonhabitual users. However, not all studies find this “tolerance effect”,[96][29] and a recent meta-analysis concludes that caffeine improves exercise performance even in habitual caffeine users.[97]
It has been suggested that athletes who regularly use caffeine and are seeking performance benefits could take a slightly higher dose before exercise.[98] Another solution would be for athletes to abstain from caffeine intake in the days before a competition to remove tolerance and thereby exploit the performance-enhancing effects of pre-exercise caffeine intake. However, this approach is not recommended because withdrawing from caffeine can increase a person's susceptibility to the unwanted side effects of caffeine.[98] Given the between-person variability in the response to caffeine (both benefits and side effects), the preferred solution is for athletes to experiment with caffeine doses and timing before competition day.[98]
While some epidemiological studies find a relationship between greater long-term daily caffeine intake (through coffee or tea) and a lower risk of later life cognitive decline or cognitive disorders, others find an inverse relationship and some studies find no relationship at all.[99][100][101][102] A main limitation in such epidemiological studies is that caffeine intake is based on a questionnaire-based retrospective recall of caffeine-containing beverages and foods overall several months or years. Furthermore, the epidemiological data finds no evidence of a dose-response relationship between long-term caffeine intake and cognitive function.[100][101] Consequently, an overall conclusion cannot be made from the epidemiological data; well-designed randomized controlled trials or cohort studies are needed to make firm conclusions about the effects of long-term caffeine intake and the risk of cognitive disorders like dementia and Alzheimer’s disease.
Caffeine is a “monitored” substance — i.e. WADA continues to monitor caffeine to detect patterns of misuse in sport — but it is not currently on the World Anti-Doping Agency’s (WADA) prohibited list for in or out-of-competition use. However, athletes should check the yearly updates to WADA’s prohibited list as well as the specific rules published by the governing body of their sport (e.g., the NCAA).
Coffee contains a mix of nutrients, including carbohydrates, fats, proteins, polyphenols and flavonoids, and, of course, caffeine.[103] While very few randomized controlled trials have explored the effect of coffee on exercise performance,[103] some studies have found a beneficial effect of pre-exercise coffee consumption on exercise performance.[104][105][106] However, examining the effect of coffee on performance creates a practical problem: The optimal caffeine dose for increasing exercise performance is 3 to 6 mg per kg bodyweight (approximately 200–400 mg in a 70 kg person) taken around 60 minutes before exercise.[1] Because a cup of coffee could contain anywhere between 50 and 200 mg of caffeine, and because the precise amount of caffeine per cup is highly variable depending on the type of bean, how the bean is roasted, and the method of preparation,[7][8] estimating the amount of caffeine in an “average” cup of coffee is difficult and has a large margin of error. While it has been suggested that 2 to 4 cups of coffee consumed around 60 minutes before exercise should improve exercise performance in most people,[37][103] in order to accurately exploit the performance-enhancing effect of caffeine, choosing a sports nutrition product with a known amount of caffeine might be preferable to drinking an “average” cup of coffee.
As an alternative to ingesting caffeine, a small number of studies have tested the effects of chewing a caffeine-containing gum or using a caffeine-containing mouth rinse. The evidence suggests that caffeine gum may have a very small beneficial effect on endurance and strength-related outcomes, but only if the gum is chewed less than 15 minutes before exercise and contains a caffeine dose of at least 3 mg per kilogram of bodyweight (equivalent to approximately 210 mg for a 70 kg person).[111] Meanwhile, the results from the small number of published mouth rinse studies are equivocal, so it is currently unclear whether rinsing the mouth with caffeine is as effective as ingesting it.[112][113] Further high-quality research is needed to clarify whether these alternative routes of caffeine delivery are effective alternatives to ingesting caffeine.
Caffeine is generally safe for most people if used within the recommended amounts (up to 400 mg per day in a healthy adult, or up to 200 mg in a single dose — see the dosage information section.[3] Furthermore, caffeine-containing beverages like tea and coffee are socially acceptable and widely used.[45][12] However, caffeine has several side effects and drawbacks.
The side effects of caffeine include a short-lived rise in heart rate and blood pressure, heart palpitations, headache, increased urine output, nervousness, gastrointestinal problems, etc.[20][21][22][23] Caffeine can also raise heart rate during and following exercise.[46][2] Some of these side effects, particularly those related to heart rate, blood pressure, and urine output, may subside with regular use due to increased tolerance.[20][22][8][13][14] Because caffeine can cause mild drug dependence, some people also experience withdrawal symptoms — headache, drowsiness, and irritability — when they stop using caffeine after regular daily intake.[47]
One of the major drawbacks is that caffeine impairs sleep quality and can reduce sleep duration.[48][49] A recent meta-analysis found that to prevent deleterious effects on subsequent sleep, coffee should be consumed at least 9 hours before bedtime, and a caffeine-containing pre-workout supplement should be consumed at least 13 hours before bedtime.[48]
Sadly, there have been several case reports of caffeine toxicity[50][51][52][53] and caffeine-related death.[54][55][56][57] However, considering the high global prevalence of caffeine intake, these cases are rare.[45][12][58] Such cases also typically involve exceptionally high caffeine intakes (usually unintentional but sometimes purposeful) or caffeine intake combined with other drugs and alcohol.[54] Some instances of toxicity and death involve energy drinks — beverages that contain as much as 300 mg of caffeine plus other stimulants like taurine, in addition to B vitamins and sugar.[59] At the population level, energy drinks are unlikely to cause harm because there is a low prevalence of consumption.[60][61] However, in certain groups (children, teenagers, people with underlying heart conditions), health risks can emerge when people far exceed recommended daily caffeine intake limits by consuming energy drinks with other caffeine-containing foods and beverages.[60][61]
Some epidemiological studies have found a relationship between high daily caffeine intake (e.g. more than 4 to 6 cups of coffee per day) and increased cardiovascular disease risk.[62][63][64][65] However, the evidence from mendelian randomization studies and meta-analyses of epidemiological studies shows that caffeine (or coffee) intake does not increase the risk of cardiovascular disease, including coronary artery disease, stroke, heart failure, and cardiovascular disease mortality,[61][65][66][67] and that moderate intake might even be protective.[68][69][70][71]
Some studies also find a relationship between caffeine intake during pregnancy and poorer health outcomes for mother and baby following birth; however, the entirety of the evidence is equivocal.[72][73] Furthermore, it is unclear whether reducing caffeine intake during pregnancy has any effect on pregnancy outcomes, including infant health and birth weight.[72][73] However, some authorities recommend that pregnant women limit their caffeine intake to 200 milligrams per day (the equivalent of 1–2 cups of coffee per day or less).[5]
Some media stories have indicated that caffeine can cause a specific, unusual heartbeat pattern (i.e., an arrhythmia), atrial fibrillation (AF). AF is a type of arrhythmia that can cause chest pain, dizziness, and fatigue. However, these media stories have been based on anecdotal case studies of individual people. When examining the data from observational research and large cohort studies, the current evidence shows that neither caffeine nor coffee is likely to increase the risk of atrial fibrillation.[107][108][109][110] However, people with heart problems should seek advice from their doctor concerning caffeine intake.
Caffeine’s mechanisms of action are not completely understood, but the proposed mechanisms likely act in unison.[14][1][74]
Caffeine’s primary action is in the central nervous system, where it prevents adenosine from binding to its receptor.[75][14][74] This affects the secretion of several neurotransmitters — norepinephrine, dopamine, acetylcholine, serotonin, GABA, etc. — involved in alertness, mood, motivation, memory, and pain perception.[14][76] For example, adenosine released during exercise decreases the secretion of dopamine, a neurotransmitter that increases mood and motivation.[1][77] So, when taken before or during exercise, caffeine allows dopamine secretion to continue, potentially maintaining mood and motivation to work hard.[77] This, coupled with lower pain perception caused by the lack of adenosine signaling, may partly explain caffeine’s sports-performance-enhancing effects.
Caffeine also plays a role in cellular calcium transport, which regulates neurotransmitter secretion in neurons and contractile force in muscle cells.[14][74] Caffeine can have a direct effect on calcium transport in muscle cells and may, therefore, have a direct effect on muscle contractile force,[1][78][79][80][81][82] providing an additional explanation for its performance-enhancing effects.
Caffeine can also inhibit a group of enzymes called cyclic nucleotide phosphodiesterases (PDEs), preventing the breakdown of cyclic adenosine monophosphate (cAMP).[14][74] This is important because cAMP stimulates the release of neurotransmitters like dopamine, epinephrine, and norepinephrine, which regulate aspects of cognitive function, including alertness, mood, motivation, and memory.[14][74] However, this mechanism requires more research and may only occur in the presence of high concentrations of caffeine, i.e., following high caffeine doses.
There are benefits associated with chronic caffeine consumption, and there are benefits associated with acute caffeine consumption that fade with tolerance; if you like the latter, cycling is mandatory. If you like the former, cycling is not needed.
Chronic non-cycled caffeine consumption (in the form of coffee and tea, both of which contain many bioactive components in addition to caffeine) is associated with a reduced risk of several diseases, including Parkinson’s disease, type 2 diabetes, and chronic liver disease.[61] Caffeine consumption has also been shown to enhance performance on attention tasks, regardless of the participants’ caffeine consumption habits.[114] Clearly, cycling isn’t needed to obtain these benefits. So why cycle? The question usually comes up in the context of exercise performance enhancement.
Supplementation with 3–6 mg of caffeine per kg of body weight before exercise has been consistently shown to enhance performance over a wide range of exercise intensities and durations, with beneficial effects on aerobic exercise performance, muscular endurance and strength, sprinting, jumping, and throwing performance.[1]
Caffeine elicits an ergogenic effect (i.e., enhances exercise performance) primarily by affecting the central nervous system. Caffeine blocks adenosine receptors in the brain, resulting in increased release of neurotransmitters such as dopamine and norepinephrine, and thus increased alertness and focus. It can also reduce pain and perceived exertion during exercise.
It seems to many habitual caffeine users that habitual use reduces, or even eliminates, the ergogenic effect of acute caffeine ingestion. The rationale is that habitual caffeine use increases the number of adenosine receptors in the brain, and as such, reduces the adenosine-blocking effect of caffeine. This has been shown in rodents,[115][116] but has yet to be studied in humans. Evidence from human studies does suggest that habitual caffeine use can blunt some aspects of the physiological response to caffeine, such as an increase in plasma epinephrine levels.[117][20]
In further support of the idea that athletes should cycle caffeine in order to maximize its ergogenic effect, some studies indicate that when individuals with low habitual caffeine intake (< 75 mg/day) consume 3 mg of caffeine per kg of body weight daily for 20–28 days, the ergogenic effect of caffeine is reduced.[93][94]
However, in a 2022 meta-analysis of 59 studies that investigated the effect of acute supplementation with caffeine on exercise performance in habitual caffeine consumers, it was found that acute supplementation with caffeine improved performance during endurance, power, and strength exercises.[97] Moreover, the performance-enhancing effects of an acute dose of caffeine were independent of whether or not that dose was greater or smaller than participants' habitual consumption, as well as being independent of sex or training status.
The results of this meta-analysis indicate that caffeine does not lose its ergogenicity with chronic use; that is, even with daily caffeine consumption, ingesting some caffeine about an hour before exercise will still have a positive impact on performance in people who benefit from caffeine (which is not everyone). But is it possible that skipping a cup of coffee today will enhance the effects of a cup of coffee on exercise performance tomorrow? Probably not.
Two separate studies reported that abstaining from caffeine for four days prior to an exercise test did not enhance the effect of acute supplementation with caffeine,[118][119] and shorter withdrawal periods (24–48 hours) don’t appear to be useful either.[97] Therefore, the available evidence does not support the common practice of utilizing a caffeine withdrawal period before an important training session or competition to maximize caffeine’s ergogenic effect. In fact, doing so may even be detrimental, as a four-day withdrawal period was found to result in a number of side effects, including headaches, fatigue, impaired focus, and a lack of motivation.
It remains to be determined whether a longer withdrawal period can boost caffeine’s ergogenic effect, but considering the fact that acute supplementation with caffeine enhances exercise performance in habitual users, and abstaining from caffeine tends to result in unfavorable symptoms, the juice is unlikely to be worth the squeeze if the purpose of a caffeine withdrawal period is solely to maximize caffeine’s ergogenic effect on a specific day.
Over the course of a day, you get sleepy as adenosine binds to A1 receptors in your brain. Caffeine blocks adenosine from binding, thus making you feel alert and also helping you feel better.
Caffeine is the most widely consumed psychoactive drug in the world, largely due to its mood-enhancing and stimulatory effects.
Despite widespread consumption, few people are actually aware of how caffeine works in the body.
We thought you might be interested in learning exactly how caffeine works in your body after you ingest it and it enters the brain.
But first: how adenosine makes you feel sleepy
The key player here is adenosine.
If you remember your high school biology, think of the lock-and-key model.
Adenosine is a key that opens up a variety of locks, with the locks actually being receptors in the brain.
Once adenosine (the key) locks into a certain receptor (the lock) in the brain, it has a unique effect on the brain.
There are a host of different receptors in your brain, so different ones have different effects. The one we’re interested in is the A1 receptor. Once adenosine locks with the A1 receptor, it promotes muscle relaxation and sleepiness, which is why people get tired as the day progresses.
Furthermore, adenosine can bind to the A2A receptor. When it binds, this interferes with the release of mood-improving neurotransmitters, such as dopamine.
Adenosine itself is produced primarily from physical work and intensive brain use. Thus, over the course of the day, your body accumulates adenosine.
If only there was something that could get in the way of adenosine from locking into the A1 receptor...
Adenosine is one of the best-known sleep-regulating molecules. Located in your central nervous system, it helps get you sleepy as the day winds down, among other effects.
What caffeine does in your brain
Before caffeine
When you first wake up, your body has metabolized away the adenosine molecules. You’re a bit groggy, but you’re waking up.
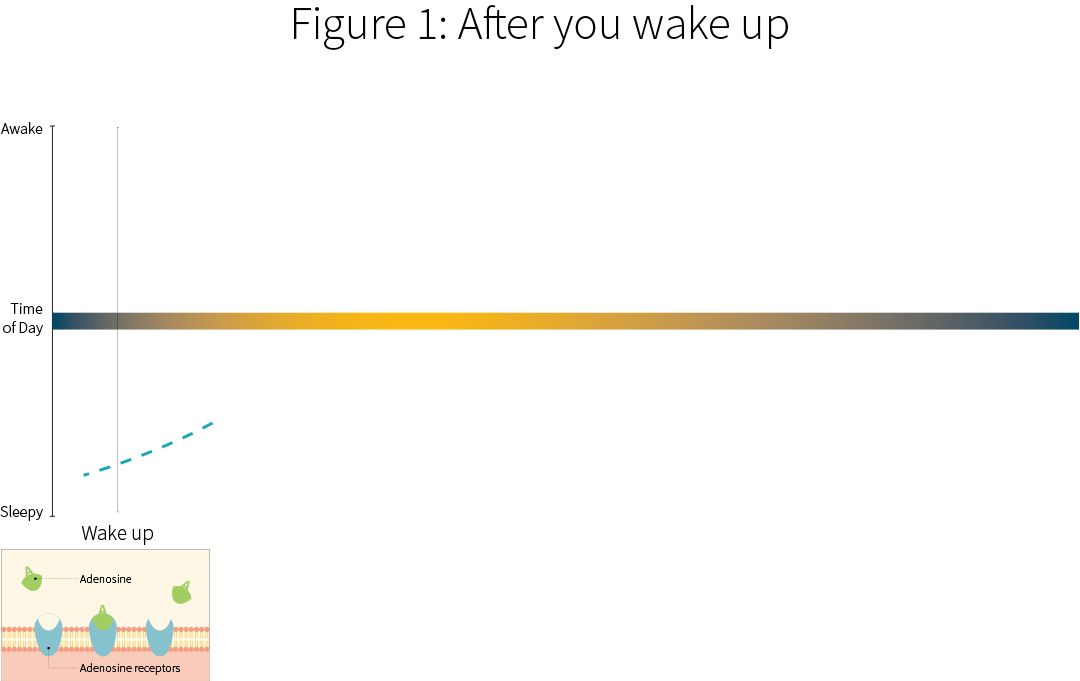
Ingesting Caffeine
Most people initially drink caffeine in the form of a beverage. It’s absorbed in the small intestines within an hour, and becomes available throughout the blood and most parts of the body, including your brain.
As it starts entering your brain, it starts competing with adenosine.

Peak Concentration
Blood concentrations of caffeine tend to peak within two hours, which also means that brain concentrations of caffeine are at their peaks. The caffeine in your brain is competing with adenosine and preventing it from binding to A1 receptors. This is what gives you a jolt of wakefulness.
To be precise, the caffeine doesn’t actually lock in with the A1 receptor. It’s more like something that gets in the way and occupies the lock, rather than actually unlocking it.
It similarly gets in the way of the A2A receptor, which can help promote the release of dopamine and glutamate (and make you feel good after you drink coffee)!

Decreasing caffeine
Eventually, caffeine molecules will unbind from the adenosine receptors (as all molecules generally do).
Most of caffeine is metabolized through the CYP1A1/2 enzymes into various substances such as araxanthine, theobromine, and theophylline.
The half-life (the amount of time it takes for the concentration of a substance to be halved) of caffeine in the body ranges from three to ten hours depending on the amount of CYP1A1 in the body, which varies from individual to individual.

Feeling sleepy again
By early evening, most of the caffeine from your morning cup of coffee has metabolized. There are significantly fewer caffeine molecules occupying the A1 receptors, so adenosine starts binding to them.
This starts promoting muscle relaxation and sleepiness, and that’s why you start feeling sleepy.

When you go to sleep and your body starts recovering, the adenosine molecules are metabolized. This is why sleep is so important - one of the issues with a lack of sleep is the increase in adenosine molecules. This then takes us back to the "Before Caffeine" step.
Of course, you can always attempt to drink a larger dose of caffeine at one sitting, or drink caffeine multiple times during the day to keep sleepiness at bay. But that’s not really a sustainable strategy.
Caffeine allows people to remain awake by competing with a molecule that promotes sleepiness called adenosine. Caffeine has a similar shape to adenosine and prevents it from binding to its receptors.
For more information, check out our in-depth caffeine page.
As an alternative to ingesting caffeine, a small number of studies have tested the effects of chewing a caffeine-containing gum or mouth-rinsing a caffeine-containing solution. The evidence suggests that caffeine gum may have a very small beneficial effect on endurance and strength-related outcomes, but only if the gum is chewed less than 15 minutes before exercise at a caffeine dose of at least 3 mg per kilogram of bodyweight (equivalent to approximately 210 mg in a 70 kg person).[111] Meanwhile, the results from the small number of published mouth rinse studies are equivocal, so it is currently unclear whether mouth-rinsing caffeine is as effective as ingesting it.[112][113] Further high-quality research is needed to clarify whether these alternative routes of caffeine delivery are a valid alternative to ingesting caffeine.
Update History
Full page update
Written By
Edited By
Reviewed By
One new meta analysis added
Reviewed By
References
- ^Nanci S Guest, Trisha A VanDusseldorp, Michael T Nelson, Jozo Grgic, Brad J Schoenfeld, Nathaniel D M Jenkins, Shawn M Arent, Jose Antonio, Jeffrey R Stout, Eric T Trexler, Abbie E Smith-Ryan, Erica R Goldstein, Douglas S Kalman, Bill I CampbellInternational society of sports nutrition position stand: caffeine and exercise performanceJ Int Soc Sports Nutr.(2021 Jan 2)
- ^de Souza JG, Del Coso J, Fonseca FS, Silva BVC, de Souza DB, da Silva Gianoni RL, Filip-Stachnik A, Serrão JC, Claudino JGRisk or benefit? Side effects of caffeine supplementation in sport: a systematic review.Eur J Nutr.(2022-Dec)
- ^Wikoff D, Welsh BT, Henderson R, Brorby GP, Britt J, Myers E, Goldberger J, Lieberman HR, O'Brien C, Peck J, Tenenbein M, Weaver C, Harvey S, Urban J, Doepker CSystematic review of the potential adverse effects of caffeine consumption in healthy adults, pregnant women, adolescents, and childrenFood Chem Toxicol.(2017 Nov)
- ^Poole R, Kennedy OJ, Roderick P, Fallowfield JA, Hayes PC, Parkes JCoffee consumption and health: umbrella review of meta-analyses of multiple health outcomesBMJ.(2017 Nov 22)
- ^EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA)Scientific Opinion on the Safety of CaffeineEFSA Journal.(May 2015)
- ^US Food and Drug AdministrationSpilling the Beans: How Much Caffeine is Too Much?FDA Consumer Updates.(2023 September)
- ^Awwad S, Issa R, Alnsour L, Albals D, Al-Momani IQuantification of Caffeine and Chlorogenic Acid in Green and Roasted Coffee Samples Using HPLC-DAD and Evaluation of the Effect of Degree of Roasting on Their Levels.Molecules.(2021-Dec-11)
- ^Heckman MA, Weil J, Gonzalez de Mejia ECaffeine (1, 3, 7-trimethylxanthine) in foods: a comprehensive review on consumption, functionality, safety, and regulatory mattersJ Food Sci.(2010 Apr)
- ^US Food and Drug AdministrationFDA Warns Consumers About Pure and Highly Concentrated Caffeine.FDA Consumer Updates.(2018 April)
- ^PubChem Compound Summary for CID 2519, CaffeineNational Center for Biotechnology Information, National Library of Medicine.(2023 Oct)
- ^PubChem Compound Summary for CID 6241, Caffeine citrateNational Center for Biotechnology Information, National Library of Medicine.(2023 Oct)
- ^Fulgoni VL, Keast DR, Lieberman HRTrends in intake and sources of caffeine in the diets of US adults: 2001-2010.Am J Clin Nutr.(2015-May)
- ^Nehlig A, Daval JL, Debry GCaffeine and the central nervous system: mechanisms of action, biochemical, metabolic and psychostimulant effects.Brain Res Brain Res Rev.(1992)
- ^Fiani B, Zhu L, Musch BL, Briceno S, Andel R, Sadeq N, Ansari AZThe Neurophysiology of Caffeine as a Central Nervous System Stimulant and the Resultant Effects on Cognitive Function.Cureus.(2021-May-14)
- ^Planning Committee for a Workshop on Potential Health Hazards Associated with Consumption of Caffeine in Food and Dietary Supplements, Food and Nutrition Board, Board on Health Sciences Policy, Institute of MedicineCaffeine in Food and Dietary Supplements: Examining Safety: Workshop Summary
- ^Harty PS, Zabriskie HA, Erickson JL, Molling PE, Kerksick CM, Jagim ARMulti-ingredient pre-workout supplements, safety implications, and performance outcomes: a brief reviewJ Int Soc Sports Nutr.(2018 Aug 8)
- ^Christopher J Derry, Sheena Derry, R Andrew MooreCaffeine as an analgesic adjuvant for acute pain in adultsCochrane Database Syst Rev.(2014 Dec 11)
- ^Derry S, Wiffen PJ, Moore RASingle dose oral ibuprofen plus caffeine for acute postoperative pain in adults.Cochrane Database Syst Rev.(2015-Jul-14)
- ^Bruschettini M, Brattström P, Russo C, Onland W, Davis PG, Soll RCaffeine dosing regimens in preterm infants with or at risk for apnea of prematurity.Cochrane Database Syst Rev.(2023-Apr-11)
- ^Robertson D, Wade D, Workman R, Woosley RL, Oates JATolerance to the humoral and hemodynamic effects of caffeine in man.J Clin Invest.(1981-Apr)
- ^Butler JM, Frampton CM, Moore G, Barclay ML, Jardine DLImmediate effect of caffeine on sympathetic nerve activity: why coffee is safe? A single-centre crossover study.Clin Auton Res.(2023-Aug-20)
- ^Maughan RJ, Griffin JCaffeine ingestion and fluid balance: a reviewJ Hum Nutr Diet.(2003 Dec)
- ^Armstrong LECaffeine, body fluid-electrolyte balance, and exercise performance.Int J Sport Nutr Exerc Metab.(2002-Jun)
- ^Lorenzo Calvo J, Fei X, Domínguez R, Pareja-Galeano HCaffeine and Cognitive Functions in Sports: A Systematic Review and Meta-Analysis.Nutrients.(2021-Mar-06)
- ^Irwin C, Khalesi S, Desbrow B, McCartney DEffects of acute caffeine consumption following sleep loss on cognitive, physical, occupational and driving performance: A systematic review and meta-analysis.Neurosci Biobehav Rev.(2020-Jan)
- ^Crawford C, Teo L, Lafferty L, Drake A, Bingham JJ, Gallon MD, O'Connell ML, Chittum HK, Arzola SM, Berry KCaffeine to optimize cognitive function for military mission-readiness: a systematic review and recommendations for the field.Nutr Rev.(2017-Jun-01)
- ^Ker K, Edwards PJ, Felix LM, Blackhall K, Roberts ICaffeine for the prevention of injuries and errors in shift workers.Cochrane Database Syst Rev.(2010-May-12)
- ^Welsh EJ, Bara A, Barley E, Cates CJCaffeine for asthma.Cochrane Database Syst Rev.(2010-Jan-20)
- ^Conger SA, Tuthill LM, Millard-Stafford MLDoes Caffeine Increase Fat Metabolism? A Systematic Review and Meta-Analysis.Int J Sport Nutr Exerc Metab.(2023-Mar-01)
- ^Collado-Mateo D, Lavín-Pérez AM, Merellano-Navarro E, Coso JDEffect of Acute Caffeine Intake on the Fat Oxidation Rate during Exercise: A Systematic Review and Meta-Analysis.Nutrients.(2020-Nov-24)
- ^Hursel R, Viechtbauer W, Dulloo AG, Tremblay A, Tappy L, Rumpler W, Westerterp-Plantenga MSThe effects of catechin rich teas and caffeine on energy expenditure and fat oxidation: a meta-analysisObes Rev.(2011 Jul)
- ^Harpaz E, Tamir S, Weinstein A, Weinstein YThe effect of caffeine on energy balance.J Basic Clin Physiol Pharmacol.(2017-Jan-01)
- ^Schubert MM, Irwin C, Seay RF, Clarke HE, Allegro D, Desbrow BCaffeine, coffee, and appetite control: a review.Int J Food Sci Nutr.(2017-Dec)
- ^Tabrizi R, Saneei P, Lankarani KB, Akbari M, Kolahdooz F, Esmaillzadeh A, Nadi-Ravandi S, Mazoochi M, Asemi ZThe effects of caffeine intake on weight loss: a systematic review and dos-response meta-analysis of randomized controlled trials.Crit Rev Food Sci Nutr.(2019)
- ^Glaister M, Gissane CCaffeine and Physiological Responses to Submaximal Exercise: A Meta-AnalysisInt J Sports Physiol Perform.(2017 Sep 5)
- ^M Doherty, P M SmithEffects of caffeine ingestion on rating of perceived exertion during and after exercise: a meta-analysisScand J Med Sci Sports.(2005 Apr)
- ^Grgic J, Grgic I, Pickering C, Schoenfeld BJ, Bishop DJ, Pedisic ZWake up and smell the coffee: caffeine supplementation and exercise performance-an umbrella review of 21 published meta-analysesBr J Sports Med.(2019 Mar 29)
- ^Wang Z, Qiu B, Gao J, Del Coso JEffects of Caffeine Intake on Endurance Running Performance and Time to Exhaustion: A Systematic Review and Meta-Analysis.Nutrients.(2022-Dec-28)
- ^Arguedas-Soley A, Townsend I, Hengist A, Betts JAcute caffeine supplementation and live match-play performance in team-sports: A systematic review (2000-2021).J Sports Sci.(2022-Apr)
- ^Salinero JJ, Lara B, Del Coso JEffects of acute ingestion of caffeine on team sports performance: a systematic review and meta-analysis.Res Sports Med.(2019)
- ^Jozo Grgic, Juan Del CosoErgogenic Effects of Acute Caffeine Intake on Muscular Endurance and Muscular Strength in Women: A Meta-AnalysisInt J Environ Res Public Health.(2021 May 27)
- ^Gomez-Bruton A, Marin-Puyalto J, Muñiz-Pardos B, Matute-Llorente A, Del Coso J, Gomez-Cabello A, Vicente-Rodriguez G, Casajus JA, Lozano-Berges GDoes Acute Caffeine Supplementation Improve Physical Performance in Female Team-Sport Athletes? Evidence from a Systematic Review and Meta-Analysis.Nutrients.(2021-Oct-19)
- ^Mielgo-Ayuso J, Marques-Jiménez D, Refoyo I, Del Coso J, León-Guereño P, Calleja-González JEffect of Caffeine Supplementation on Sports Performance Based on Differences Between Sexes: A Systematic ReviewNutrients.(2019 Sep 30)
- ^Maughan RJ, Burke LM, Dvorak J, Larson-Meyer DE, Peeling P, Phillips SM, Rawson ES, Walsh NP, Garthe I, Geyer H, Meeusen R, van Loon LJC, Shirreffs SM, Spriet LL, Stuart M, Vernec A, Currell K, Ali VM, Budgett RG, Ljungqvist A, Mountjoy M, Pitsiladis YP, Soligard T, Erdener U, Engebretsen LIOC consensus statement: dietary supplements and the high-performance athleteBr J Sports Med.(2018 Apr)
- ^Reyes CM, Cornelis MCCaffeine in the Diet: Country-Level Consumption and Guidelines.Nutrients.(2018-Nov-15)
- ^Benjamim CJR, Kliszczewicz B, Garner DM, Cavalcante TCF, da Silva AAM, Santana MDR, Valenti VEIs Caffeine Recommended Before Exercise? A Systematic Review To Investigate Its Impact On Cardiac Autonomic Control Via Heart Rate And Its Variability.J Am Coll Nutr.(2020-08)
- ^Juliano LM, Griffiths RRA critical review of caffeine withdrawal: empirical validation of symptoms and signs, incidence, severity, and associated featuresPsychopharmacology (Berl).(2004 Oct)
- ^Gardiner C, Weakley J, Burke LM, Roach GD, Sargent C, Maniar N, Townshend A, Halson SLThe effect of caffeine on subsequent sleep: A systematic review and meta-analysis.Sleep Med Rev.(2023-Jun)
- ^Clark I, Landolt HPCoffee, caffeine, and sleep: A systematic review of epidemiological studies and randomized controlled trialsSleep Med Rev.(2017 Feb)
- ^Nakaoka S, Kawasaki Y, Inomata S, Makimoto M, Yoshida TCaffeine Toxicity in a Preterm Neonate.Pediatr Neonatol.(2017-Aug)
- ^Ehab HamedCaffeine Toxicity Following Ingestion of an Exercise Supplement by a Patient with Type 1 DiabetesEur J Case Rep Intern Med.(2018 Oct 24)
- ^Willson CThe clinical toxicology of caffeine: A review and case studyToxicol Rep.(2018 Nov 3)
- ^Tsuyoshi Nojima, Hiromichi Naito, Yoshinori Kosaki, Takaaki Osako, Kimiaki Tanaka, Atsuo Murata, Atsunori NakaoCaffeine Intoxication in Pregnancy; a case ReportArch Acad Emerg Med.(2019 Nov 16)
- ^Jones AWReview of Caffeine-Related Fatalities along with Postmortem Blood Concentrations in 51 Poisoning DeathsJ Anal Toxicol.(2017 Apr 1)
- ^Grémain V, Chevillard L, Saussereau E, Schnell G, Mégarbane BMassive suicidal ingestion of caffeine: a case report with investigation of the cardiovascular effect/concentration relationships.Clin Toxicol (Phila).(2021-Oct)
- ^Jabbar SB, Hanly MGFatal caffeine overdose: a case report and review of literatureAm J Forensic Med Pathol.(2013 Dec)
- ^Winek CL, Wahba W, Williams K, Blenko J, Janssen JCaffeine fatality: a case report.Forensic Sci Int.(1985)
- ^Temple JL, Bernard C, Lipshultz SE, Czachor JD, Westphal JA, Mestre MAThe Safety of Ingested Caffeine: A Comprehensive ReviewFront Psychiatry.(2017 May 26)
- ^Jagim AR, Harty PS, Fischer KM, Kerksick CM, Erickson JLAdverse Events Reported to the United States Food and Drug Administration Related to Caffeine-Containing Products.Mayo Clin Proc.(2020-Aug)
- ^Thomson BM, Campbell DM, Cressey P, Egan U, Horn BEnergy drink consumption and impact on caffeine risk.Food Addit Contam Part A Chem Anal Control Expo Risk Assess.(2014)
- ^van Dam RM, Hu FB, Willett WCCoffee, Caffeine, and Health.N Engl J Med.(2020-07-23)
- ^Zheng H, Lin F, Xin N, Yang L, Zhu PAssociation of Coffee, Tea, and Caffeine Consumption With All-Cause Risk and Specific Mortality for Cardiovascular Disease Patients.Front Nutr.(2022)
- ^Zhou A, Hyppönen ELong-term coffee consumption, caffeine metabolism genetics, and risk of cardiovascular disease: a prospective analysis of up to 347,077 individuals and 8368 casesAm J Clin Nutr.(2019 Mar 1)
- ^Grioni S, Agnoli C, Sieri S, Pala V, Ricceri F, Masala G, Saieva C, Panico S, Mattiello A, Chiodini P, Tumino R, Frasca G, Iacoviello L, de Curtis A, Vineis P, Krogh VEspresso coffee consumption and risk of coronary heart disease in a large Italian cohort.PLoS One.(2015)
- ^Said MA, van de Vegte YJ, Verweij N, van der Harst PAssociations of Observational and Genetically Determined Caffeine Intake With Coronary Artery Disease and Diabetes Mellitus.J Am Heart Assoc.(2020-Dec-15)
- ^Park Y, Cho H, Myung SKEffect of Coffee Consumption on Risk of Coronary Heart Disease in a Systematic Review and Meta-Analysis of Prospective Cohort Studies.Am J Cardiol.(2023-Jan-01)
- ^Qian Y, Ye D, Huang H, Wu DJH, Zhuang Y, Jiang X, Mao YCoffee Consumption and Risk of Stroke: A Mendelian Randomization Study.Ann Neurol.(2020-Apr)
- ^Ming Ding, Shilpa N Bhupathiraju, Ambika Satija, Rob M van Dam, Frank B HuLong-term coffee consumption and risk of cardiovascular disease: a systematic review and a dose-response meta-analysis of prospective cohort studiesCirculation.(2014 Feb 11)
- ^Alessio Crippa, Andrea Discacciati, Susanna C Larsson, Alicja Wolk, Nicola OrsiniCoffee consumption and mortality from all causes, cardiovascular disease, and cancer: a dose-response meta-analysisAm J Epidemiol.(2014 Oct 15)
- ^Susanna C Larsson, Nicola OrsiniCoffee consumption and risk of stroke: a dose-response meta-analysis of prospective studiesAm J Epidemiol.(2011 Nov 1)
- ^Shao C, Tang H, Wang X, He JCoffee Consumption and Stroke Risk: Evidence from a Systematic Review and Meta-Analysis of more than 2.4 Million Men and Women.J Stroke Cerebrovasc Dis.(2021-Jan)
- ^ACOG CommitteeOpinion No. 462: Moderate caffeine consumption during pregnancy.Obstet Gynecol.(2010 Aug)
- ^Shayesteh Jahanfar, Sharifah Halimah JaafarEffects of restricted caffeine intake by mother on fetal, neonatal and pregnancy outcomesCochrane Database Syst Rev.(2015 Jun 9)
- ^Simone Cappelletti, Daria Piacentino, Gabriele Sani, Mariarosaria AromatarioCaffeine: cognitive and physical performance enhancer or psychoactive drug?Curr Neuropharmacol.(2015 Jan)
- ^Kenneth A Jacobson, Zhan-Guo Gao, Pierre Matricon, Matthew T Eddy, Jens CarlssonAdenosine A2A receptor antagonists: from caffeine to selective non-xanthinesBr J Pharmacol.(2022 Jul)
- ^Haddad M, Cherchi F, Alsalem M, Al-Saraireh YM, Madae'en SAdenosine Receptors as Potential Therapeutic Analgesic Targets.Int J Mol Sci.(2023-Aug-24)
- ^Roelands B, Meeusen RAlterations in central fatigue by pharmacological manipulations of neurotransmitters in normal and high ambient temperature.Sports Med.(2010-Mar-01)
- ^Black CD, Waddell DE, Gonglach ARCaffeine's Ergogenic Effects on Cycling: Neuromuscular and Perceptual Factors.Med Sci Sports Exerc.(2015-Jun)
- ^Rousseau E, Ladine J, Liu QY, Meissner GActivation of the Ca2+ release channel of skeletal muscle sarcoplasmic reticulum by caffeine and related compounds.Arch Biochem Biophys.(1988-Nov-15)
- ^Tarnopolsky M, Cupido CCaffeine potentiates low frequency skeletal muscle force in habitual and nonhabitual caffeine consumersJ Appl Physiol (1985).(2000 Nov)
- ^Domaszewski P, Pakosz P, Konieczny M, Bączkowicz D, Sadowska-Krępa ECaffeine-Induced Effects on Human Skeletal Muscle Contraction Time and Maximal Displacement Measured by Tensiomyography.Nutrients.(2021-Mar-02)
- ^Cristina-Souza G, Santos PS, Santos-Mariano AC, Coelho DB, Rodacki A, DE-Oliveira FR, Bishop DJ, Bertuzzi R, Lima-Silva AECaffeine Increases Endurance Performance via Changes in Neural and Muscular Determinants of Performance Fatigability.Med Sci Sports Exerc.(2022-Sep-01)
- ^Fulton JL, Dinas PC, Carrillo AE, Edsall JR, Ryan EJ, Ryan EJImpact of Genetic Variability on Physiological Responses to Caffeine in Humans: A Systematic ReviewNutrients.(2018 Sep 25)
- ^Cornelis MC, Monda KL, Yu K, Paynter N, Azzato EM, Bennett SN, Berndt SI, Boerwinkle E, Chanock S, Chatterjee N, Couper D, Curhan G, Heiss G, Hu FB, Hunter DJ, Jacobs K, Jensen MK, Kraft P, Landi MT, Nettleton JA, Purdue MP, Rajaraman P, Rimm EB, Rose LM, Rothman N, Silverman D, Stolzenberg-Solomon R, Subar A, Yeager M, Chasman DI, van Dam RM, Caporaso NEGenome-wide meta-analysis identifies regions on 7p21 (AHR) and 15q24 (CYP1A2) as determinants of habitual caffeine consumption.PLoS Genet.(2011-Apr)
- ^Kapellou A, King A, Graham CAM, Pilic L, Mavrommatis YGenetics of caffeine and brain-related outcomes - a systematic review of observational studies and randomized trials.Nutr Rev.(2023-Apr-08)
- ^Jozo Grgic, Craig Pickering, Juan Del Coso, Brad J Schoenfeld, Pavle MikulicCYP1A2 genotype and acute ergogenic effects of caffeine intake on exercise performance: a systematic reviewEur J Nutr.(2020 Nov 2)
- ^Barreto G, Esteves GP, Marticorena F, Oliveira TN, Grgic J, Saunders BCaffeine, CYP1A2 Genotype and Exercise Performance: A Systematic Review and Meta-analysis.Med Sci Sports Exerc.(2023-Oct-12)
- ^Mégane Erblang, Catherine Drogou, Danielle Gomez-Merino, Arnaud Metlaine, Anne Boland, Jean François Deleuze, Claire Thomas, Fabien Sauvet, Mounir ChennaouiThe Impact of Genetic Variations in ADORA2A in the Association between Caffeine Consumption and SleepGenes (Basel).(2019 Dec 6)
- ^Rafael Almeida Nunes, Diego Robles Mazzotti, Camila Hirotsu, Monica L Andersen, Sergio Tufik, Lia BittencourtThe association between caffeine consumption and objective sleep variables is dependent on ADORA2A c.1083T>C genotypesSleep Med.(2017 Feb)
- ^Byrne EM, Johnson J, McRae AF, Nyholt DR, Medland SE, Gehrman PR, Heath AC, Madden PA, Montgomery GW, Chenevix-Trench G, Martin NGA genome-wide association study of caffeine-related sleep disturbance: confirmation of a role for a common variant in the adenosine receptor.Sleep.(2012-Jul-01)
- ^Virgili J, Motitis P, Julal G, Mavrommatis Y, Pilic LThe impact of genetic variability on the relationship between caffeine and cardiometabolic outcomes: A systematic review.Nutr Bull.(2023-Mar)
- ^J Bangsbo, K Jacobsen, N Nordberg, N J Christensen, T GrahamAcute and habitual caffeine ingestion and metabolic responses to steady-state exerciseJ Appl Physiol (1985).(1992 Apr)
- ^Ross Beaumont, Philip Cordery, Mark Funnell, Stephen Mears, Lewis James, Phillip WatsonChronic ingestion of a low dose of caffeine induces tolerance to the performance benefits of caffeineJ Sports Sci.(2017 Oct)
- ^Lara B, Ruiz-Moreno C, Salinero JJ, Del Coso JTime course of tolerance to the performance benefits of caffeinePLoS One.(2019 Jan 23)
- ^Bell DG, McLellan TMExercise endurance 1, 3, and 6 h after caffeine ingestion in caffeine users and nonusersJ Appl Physiol (1985).(2002 Oct)
- ^Gonçalves LS, Painelli VS, Yamaguchi G, de Oliveira LF, Saunders B, da Silva RP, Maciel E, Artioli GG, Roschel H, Gualano BDispelling the myth that habitual caffeine consumption influences the performance response to acute caffeine supplementationJ Appl Physiol (1985).(2017 May 11)
- ^Arthur Carvalho, Felipe Miguel Marticorena, Beatriz Helena Grecco, Gabriel Barreto, Bryan SaundersCan I Have My Coffee and Drink It? A Systematic Review and Meta-analysis to Determine Whether Habitual Caffeine Consumption Affects the Ergogenic Effect of CaffeineSports Med.(2022 May 10)
- ^Pickering C, Kiely JWhat Should We Do About Habitual Caffeine Use in Athletes?Sports Med.(2019 Jun)
- ^J Q Alida Chen, Philip Scheltens, Colin Groot, Rik OssenkoppeleAssociations Between Caffeine Consumption, Cognitive Decline, and Dementia: A Systematic ReviewJ Alzheimers Dis.(2020)
- ^Panza F, Solfrizzi V, Barulli MR, Bonfiglio C, Guerra V, Osella A, Seripa D, Sabbà C, Pilotto A, Logroscino GCoffee, tea, and caffeine consumption and prevention of late-life cognitive decline and dementia: a systematic review.J Nutr Health Aging.(2015-Mar)
- ^Qing-Ping Liu, Yan-Feng Wu, Hong-Yu Cheng, Tao Xia, Hong Ding, Hui Wang, Ze-Mu Wang, Yun XuHabitual coffee consumption and risk of cognitive decline/dementia: A systematic review and meta-analysis of prospective cohort studiesNutrition.(2016 Jun)
- ^Kim YS, Kwak SM, Myung SKCaffeine intake from coffee or tea and cognitive disorders: a meta-analysis of observational studies.Neuroepidemiology.(2015)
- ^Lowery LM, Anderson DE, Scanlon KF, Stack A, Escalante G, Campbell SC, Kerksick CM, Nelson MT, Ziegenfuss TN, VanDusseldorp TA, Kalman DS, Campbell BI, Kreider RB, Antonio JInternational society of sports nutrition position stand: coffee and sports performance.J Int Soc Sports Nutr.(2023-Dec)
- ^Hodgson AB, Randell RK, Jeukendrup AEThe metabolic and performance effects of caffeine compared to coffee during endurance exercisePLoS One.(2013)
- ^Trexler ET, Smith-Ryan AE, Roelofs EJ, Hirsch KR, Mock MGEffects of coffee and caffeine anhydrous on strength and sprint performanceEur J Sport Sci.(2016 Sep)
- ^Richardson DL, Clarke NDEffect of Coffee and Caffeine Ingestion on Resistance Exercise PerformanceJ Strength Cond Res.(2016 Oct)
- ^Chayakrit Krittanawong, Anusith Tunhasiriwet, Zhen Wang, Ann M Farrell, Sakkarin Chirapongsathorn, HongJu Zhang, Takeshi Kitai, Davendra MehtaIs caffeine or coffee consumption a risk for new-onset atrial fibrillation? A systematic review and meta-analysisEur J Prev Cardiol.(2021 Oct 13)
- ^Abdelfattah R, Kamran H, Lazar J, Kassotis JDoes Caffeine Consumption Increase the Risk of New-Onset Atrial Fibrillation?Cardiology.(2018)
- ^Cheng M, Hu Z, Lu X, Huang J, Gu DCaffeine intake and atrial fibrillation incidence: dose response meta-analysis of prospective cohort studies.Can J Cardiol.(2014-Apr)
- ^Caldeira D, Martins C, Alves LB, Pereira H, Ferreira JJ, Costa JCaffeine does not increase the risk of atrial fibrillation: a systematic review and meta-analysis of observational studies.Heart.(2013-Oct)
- ^Barreto G, Loureiro LMR, Reis CEG, Saunders BEffects of caffeine chewing gum supplementation on exercise performance: A systematic review and meta-analysis.Eur J Sport Sci.(2023-May)
- ^Widemar Ferraz da Silva, João Paulo Lopes-Silva, Leandro José Camati Felippe, Guilherme Assunção Ferreira, Adriano Eduardo Lima-Silva, Marcos David Silva-CavalcanteIs caffeine mouth rinsing an effective strategy to improve physical and cognitive performance? A systematic reviewCrit Rev Food Sci Nutr.(2021 Jul 19)
- ^Ehlert AM, Twiddy HM, Wilson PBThe Effects of Caffeine Mouth Rinsing on Exercise Performance: A Systematic Review.Int J Sport Nutr Exerc Metab.(2020-Jul-15)
- ^Einöther SJ, Giesbrecht TCaffeine as an attention enhancer: reviewing existing assumptions.Psychopharmacology (Berl).(2013-Jan)
- ^Fredholm BBAdenosine actions and adenosine receptors after 1 week treatment with caffeine.Acta Physiol Scand.(1982-Jun)
- ^Fastbom J, Fredholm BBEffects of long-term theophylline treatment on adenosine A1-receptors in rat brain: autoradiographic evidence for increased receptor number and altered coupling to G-proteins.Brain Res.(1990-Jan-22)
- ^Van Soeren MH, Sathasivam P, Spriet LL, Graham TECaffeine metabolism and epinephrine responses during exercise in users and nonusers.J Appl Physiol (1985).(1993-Aug)
- ^Irwin C, Desbrow B, Ellis A, O'Keeffe B, Grant G, Leveritt MCaffeine withdrawal and high-intensity endurance cycling performanceJ Sports Sci.(2011 Mar)
- ^Van Soeren MH, Graham TEEffect of caffeine on metabolism, exercise endurance, and catecholamine responses after withdrawal.J Appl Physiol (1985).(1998-Oct)
Examine Database References
- Anaerobic Capacity - Lamina S, Musa DIErgogenic effect of varied doses of coffee-caffeine on maximal aerobic power of young African subjectsAfr Health Sci.(2009 Dec)
- Subjective Well-Being - Duncan MJ, Oxford SWThe effect of caffeine ingestion on mood state and bench press performance to failureJ Strength Cond Res.(2011 Jan)
- Blood Lactate (Exercise) - Karapetian GK, Engels HJ, Gretebeck KA, Gretebeck RJEffect of Caffeine on LT, VT and HRVTInt J Sports Med.(2012 Apr 12)
- Blood Lactate (Exercise) - Norager CB, Jensen MB, Weimann A, Madsen MRMetabolic effects of caffeine ingestion and physical work in 75-year old citizens. A randomized, double-blind, placebo-controlled, cross-over studyClin Endocrinol (Oxf).(2006 Aug)
- Heart Rate - Del Coso J, Salinero JJ, Gonzalez-Millan C, Abian-Vicen J, Perez-Gonzalez BDose response effects of a caffeine-containing energy drink on muscle performance: a repeated measures designJ Int Soc Sports Nutr.(2012 May 8)
- Training Volume - Cook C, Beaven CM, Kilduff LP, Drawer SAcute caffeine ingestion increases voluntarily chosen resistance training load following limited sleepInt J Sport Nutr Exerc Metab.(2012 Feb 15)
- Adrenaline - Anderson DE, Hickey MSEffects of caffeine on the metabolic and catecholamine responses to exercise in 5 and 28 degrees CMed Sci Sports Exerc.(1994 Apr)
- Power Output - Mora-Rodríguez R, Pallarés JG, López-Samanes A, Ortega JF, Fernández-Elías VECaffeine ingestion reverses the circadian rhythm effects on neuromuscular performance in highly resistance-trained menPLoS One.(2012)
- Power Output - Todd A Astorino, Michael N Terzi, Daniel W Roberson, Timothy R BurnettEffect of two doses of caffeine on muscular function during isokinetic exerciseMed Sci Sports Exerc.(2010 Dec)
- Power Output - Beaven CM, Hopkins WG, Hansen KT, Wood MR, Cronin JB, Lowe TEDose effect of caffeine on testosterone and cortisol responses to resistance exerciseInt J Sport Nutr Exerc Metab.(2008 Apr)
- Rate of Perceived Exertion - Bellar D, Kamimori GH, Glickman ELThe effects of low-dose caffeine on perceived pain during a grip to exhaustion taskJ Strength Cond Res.(2011 May)
- Thermogenesis - Kim TW, Shin YO, Lee JB, Min YK, Yang HMCaffeine increases sweating sensitivity via changes in sudomotor activity during physical loadingJ Med Food.(2011 Nov)
- Blood Pressure - Wedick NM, Brennan AM, Sun Q, Hu FB, Mantzoros CS, van Dam RMEffects of caffeinated and decaffeinated coffee on biological risk factors for type 2 diabetes: a randomized controlled trialNutr J.(2011 Sep 13)
- Blood Lactate (Exercise) - Glaister M, Patterson SD, Foley P, Pedlar CR, Pattison JR, McInnes GCaffeine and sprinting performance: dose responses and efficacyJ Strength Cond Res.(2012 Apr)
- Anaerobic Capacity - Schneiker KT, Bishop D, Dawson B, Hackett LPEffects of caffeine on prolonged intermittent-sprint ability in team-sport athletesMed Sci Sports Exerc.(2006 Mar)
- Anaerobic Capacity - Paton CD, Lowe T, Irvine ACaffeinated chewing gum increases repeated sprint performance and augments increases in testosterone in competitive cyclistsEur J Appl Physiol.(2010 Dec)
- Heart Rate - Desbrow B, Biddulph C, Devlin B, Grant GD, Anoopkumar-Dukie S, Leveritt MDThe effects of different doses of caffeine on endurance cycling time trial performanceJ Sports Sci.(2012)
- Aerobic Exercise Metrics - Ganio MS, Johnson EC, Klau JF, Anderson JM, Casa DJ, Maresh CM, Volek JS, Armstrong LEEffect of ambient temperature on caffeine ergogenicity during endurance exerciseEur J Appl Physiol.(2011 Jun)
- Aerobic Exercise Metrics - Womack CJ, Saunders MJ, Bechtel MK, Bolton DJ, Martin M, Luden ND, Dunham W, Hancock MThe influence of a CYP1A2 polymorphism on the ergogenic effects of caffeineJ Int Soc Sports Nutr.(2012 Mar 15)
- Rate of Perceived Exertion - Backhouse SH, Biddle SJ, Bishop NC, Williams CCaffeine ingestion, affect and perceived exertion during prolonged cyclingAppetite.(2011 Aug)
- Rate of Perceived Exertion - Astorino TA, Terzi MN, Roberson DW, Burnett TREffect of caffeine intake on pain perception during high-intensity exerciseInt J Sport Nutr Exerc Metab.(2011 Feb)
- Joint Pain (General) - Christopher J Derry, Sheena Derry, R Andrew MooreCaffeine as an analgesic adjuvant for acute pain in adultsCochrane Database Syst Rev.(2014 Dec 11)
- Memory - Smillie LD, Gökçen ECaffeine enhances working memory for extravertsBiol Psychol.(2010 Dec)
- Memory - Mednick SC, Cai DJ, Kanady J, Drummond SPComparing the benefits of caffeine, naps and placebo on verbal, motor and perceptual memoryBehav Brain Res.(2008 Nov 3)
- Memory - Childs E, de Wit HSubjective, behavioral, and physiological effects of acute caffeine in light, nondependent caffeine usersPsychopharmacology (Berl).(2006 May)
- Memory - Adan A, Serra-Grabulosa JMEffects of caffeine and glucose, alone and combined, on cognitive performanceHum Psychopharmacol.(2010 Jun)
- Memory - Ker K, Edwards PJ, Felix LM, Blackhall K, Roberts ICaffeine for the prevention of injuries and errors in shift workers.Cochrane Database Syst Rev.(2010-May-12)
- Reaction Time - Duvnjak-Zaknich DM, Dawson BT, Wallman KE, Henry GEffect of caffeine on reactive agility time when fresh and fatiguedMed Sci Sports Exerc.(2011 Aug)
- Reaction Time - Irwin C, Khalesi S, Desbrow B, McCartney DEffects of acute caffeine consumption following sleep loss on cognitive, physical, occupational and driving performance: A systematic review and meta-analysis.Neurosci Biobehav Rev.(2020-Jan)
- Reaction Time - Lorenzo Calvo J, Fei X, Domínguez R, Pareja-Galeano HCaffeine and Cognitive Functions in Sports: A Systematic Review and Meta-Analysis.Nutrients.(2021-Mar-06)
- Visual acuity (VA) - Gillingham R, Keefe AA, Keillor J, Tikuisis PEffect of caffeine on target detection and rifle marksmanshipErgonomics.(2003 Dec 15)
- Anxiety Symptoms - Rogers PJ, Hohoff C, Heatherley SV, Mullings EL, Maxfield PJ, Evershed RP, Deckert J, Nutt DJAssociation of the anxiogenic and alerting effects of caffeine with ADORA2A and ADORA1 polymorphisms and habitual level of caffeine consumptionNeuropsychopharmacology.(2010 Aug)
- Blood Lactate (Exercise) - Astrup A, Toubro S, Cannon S, Hein P, Breum L, Madsen JCaffeine: a double-blind, placebo-controlled study of its thermogenic, metabolic, and cardiovascular effects in healthy volunteersAm J Clin Nutr.(1990 May)
- Adrenaline - Keijzers GB, De Galan BE, Tack CJ, Smits PCaffeine can decrease insulin sensitivity in humansDiabetes Care.(2002 Feb)
- Insulin - Pizziol A, Tikhonoff V, Paleari CD, Russo E, Mazza A, Ginocchio G, Onesto C, Pavan L, Casiglia E, Pessina ACEffects of caffeine on glucose tolerance: a placebo-controlled studyEur J Clin Nutr.(1998 Nov)
- Glycemic Control - Greer F, Hudson R, Ross R, Graham TCaffeine ingestion decreases glucose disposal during a hyperinsulinemic-euglycemic clamp in sedentary humansDiabetes.(2001 Oct)
- Cortisol - Gavrieli A, Yannakoulia M, Fragopoulou E, Margaritopoulos D, Chamberland JP, Kaisari P, Kavouras SA, Mantzoros CSCaffeinated coffee does not acutely affect energy intake, appetite, or inflammation but prevents serum cortisol concentrations from falling in healthy menJ Nutr.(2011 Apr 1)
- Anaerobic Capacity - Hadjicharalambous M, Georgiades E, Kilduff LP, Turner AP, Tsofliou F, Pitsiladis YPInfluence of caffeine on perception of effort, metabolism and exercise performance following a high-fat mealJ Sports Sci.(2006 Aug)
- Heart Rate - Bloomer RJ, Harvey IC, Farney TM, Bell ZW, Canale REEffects of 1,3-dimethylamylamine and caffeine alone or in combination on heart rate and blood pressure in healthy men and womenPhys Sportsmed.(2011 Sep)
- Blood Flow - Shechter M, Shalmon G, Scheinowitz M, Koren-Morag N, Feinberg MS, Harats D, Sela BA, Sharabi Y, Chouraqui PImpact of acute caffeine ingestion on endothelial function in subjects with and without coronary artery diseaseAm J Cardiol.(2011 May 1)
- Wakefulness - Barry RJ, Clarke AR, Johnstone SJCaffeine and opening the eyes have additive effects on resting arousal measuresClin Neurophysiol.(2011 Oct)
- Reaction Time - Hunt MG, Momjian AJ, Wong KKEffects of diurnal variation and caffeine consumption on Test of Variables of Attention (TOVA) performance in healthy young adultsPsychol Assess.(2011 Mar)
- Heart Rate - Glaister M, Howatson G, Abraham CS, Lockey RA, Goodwin JE, Foley P, McInnes GCaffeine supplementation and multiple sprint running performanceMed Sci Sports Exerc.(2008 Oct)
- Reaction Time - Pontifex KJ, Wallman KE, Dawson BT, Goodman CEffects of caffeine on repeated sprint ability, reactive agility time, sleep and next day performanceJ Sports Med Phys Fitness.(2010 Dec)
- Running Speed - Wang Z, Qiu B, Gao J, Del Coso JEffects of Caffeine Intake on Endurance Running Performance and Time to Exhaustion: A Systematic Review and Meta-Analysis.Nutrients.(2022-Dec-28)
- Blood Pressure - Renda G, Zimarino M, Antonucci I, Tatasciore A, Ruggieri B, Bucciarelli T, Prontera T, Stuppia L, De Caterina RGenetic determinants of blood pressure responses to caffeine drinkingAm J Clin Nutr.(2012 Jan)
- Metabolic Rate - Hursel R, Viechtbauer W, Dulloo AG, Tremblay A, Tappy L, Rumpler W, Westerterp-Plantenga MSThe effects of catechin rich teas and caffeine on energy expenditure and fat oxidation: a meta-analysisObes Rev.(2011 Jul)
- Fat Oxidation - Collado-Mateo D, Lavín-Pérez AM, Merellano-Navarro E, Coso JDEffect of Acute Caffeine Intake on the Fat Oxidation Rate during Exercise: A Systematic Review and Meta-Analysis.Nutrients.(2020-Nov-24)
- Fat Oxidation - Conger SA, Tuthill LM, Millard-Stafford MLDoes Caffeine Increase Fat Metabolism? A Systematic Review and Meta-Analysis.Int J Sport Nutr Exerc Metab.(2023-Mar-01)
- Heart Rate - Glaister M, Gissane CCaffeine and Physiological Responses to Submaximal Exercise: A Meta-AnalysisInt J Sports Physiol Perform.(2017 Sep 5)
- Aerobic Exercise Metrics - Grgic J, Grgic I, Pickering C, Schoenfeld BJ, Bishop DJ, Pedisic ZWake up and smell the coffee: caffeine supplementation and exercise performance-an umbrella review of 21 published meta-analysesBr J Sports Med.(2019 Mar 29)
- Jump Height - Salinero JJ, Lara B, Del Coso JEffects of acute ingestion of caffeine on team sports performance: a systematic review and meta-analysis.Res Sports Med.(2019)
- Rate of Perceived Exertion - M Doherty, P M SmithEffects of caffeine ingestion on rating of perceived exertion during and after exercise: a meta-analysisScand J Med Sci Sports.(2005 Apr)
- Anaerobic Capacity - Carr AJ, Gore CJ, Dawson BInduced alkalosis and caffeine supplementation: effects on 2,000-m rowing performanceInt J Sport Nutr Exerc Metab.(2011 Oct)
- Asthma Symptoms - Welsh EJ, Bara A, Barley E, Cates CJCaffeine for asthma.Cochrane Database Syst Rev.(2010-Jan-20)
- Sleep Duration - Gardiner C, Weakley J, Burke LM, Roach GD, Sargent C, Maniar N, Townshend A, Halson SLThe effect of caffeine on subsequent sleep: A systematic review and meta-analysis.Sleep Med Rev.(2023-Jun)