










2023-12-18 00:30:04
Full page update
major
We updated this page and it's database with the newest meta-analyses.
Caffeine is a central nervous system stimulant that can improve sports performance and aspects of cognitive function. Caffeine or caffeine-containing beverages are often taken to help restore alertness and wakefulness or to reduce drowsiness. Despite its side effects, caffeine is safe for most people, and caffeine-containing beverages like tea and coffee are socially acceptable and widely used.
Caffeine is most often used for
Last Updated:December 18, 2023
Caffeine is a psychoactive drug that exerts a powerful stimulant effect on the central nervous system. It is naturally found in coffee, tea, and chocolate, but is also added to some soft drinks (e.g. colas), energy drinks and energy shots.[8] The National Health and Nutrition Examination Survey (NHANES) found that roughly 89% of adults in the US consume caffeine daily.[12] People often use caffeine-containing beverages to help restore alertness and wakefulness or to reduce drowsiness when feeling mentally fatigued.[13][14] Caffeine is also available as a purified powder, and it is found in some dietary supplements claiming to cause weight loss or increase energy (e.g. pre-workouts).[8][15][16] Additionally, caffeine is found in some over-the-counter drugs used to relieve pain[17][18] and in prescription drugs used to treat breathing problems in premature babies.[19]
After ingestion, caffeine acts rapidly on the central nervous system, triggering several short-lived effects including an increase in heart rate, blood pressure, and urine output.[20][21][22][23] However, with regular daily use, these symptoms of caffeine typically become less pronounced.[20][22][8][13][14]
Caffeine restores alertness and wakefulness, and reduces drowsiness during mental fatigue.[24][25][26] It also improves several aspects of cognitive function — accuracy, reasoning, memory, reaction time, attention, etc. — during cognitively and/or physically demanding tasks.[27][24][25][26] Furthermore, caffeine can improve some aspects of cognitive function that have been impaired by acute sleep deprivation.[25][26]
Caffeine can also help with pain relief. For example, when caffeine is added to analgesic (pain relieving) drugs like acetaminophen (also known as paracetamol) or ibuprofen, there is a small but clinically meaningful improvement in pain.[17][18]
In hospitals, caffeine (typically caffeine citrate) is used to treat apnea (temporary cessation of breathing) in premature babies, but both the optimal dosing strategy and caffeine’s effect on survival and subsequent neurocognitive development are unclear.[19] Furthermore, in people with asthma, caffeine may have a small beneficial effect on some aspects of respiratory function (forced expiratory volume in one minute, FEV1, and mid-expiratory flow rate) for up to four hours.[28] However, this effect also has implications for asthma diagnostics because prior caffeine ingestion may reduce the accuracy of lung function tests.[28]
Caffeine can raise whole-body fat oxidation rates, both at rest and during exercise,[29][30] and increase daily energy expenditure,[31][32] but these effects are negligible — approximately 400 kilojoules or 100 kilocalories per day. The effect of caffeine on appetite regulation and energy intake is less clear and varies depending on factors like timing and dose.[32][33] Consequently, the role of caffeine in weight loss and/or weight management is unclear. Despite one meta-analysis concluding that caffeine can promote reductions in weight, BMI, and body fat,[34] its utility is limited because the included trials used energy-restriction diets and/or ephedrine, a powerful stimulant that causes weight loss.
Caffeine is widely used in sports because it can improve both aerobic and anaerobic performance in trained and untrained individuals.[1] For example, when taken before or during exercise, caffeine has small to moderate-sized effects of lowering the rating of perceived exertion (RPE)[35][36] and improving aerobic endurance, anaerobic power, sprint speed, muscle endurance, muscle strength, muscle power (jump height), and agility.[1][37][38] These effects are found across a range of sports (running, cycling, and swimming, etc.) including skill-based team sports (e.g. basketball, soccer, etc.).[1][39][40] Furthermore, when taken before and/or during exercise, caffeine may also improve cognitive functions, such as attention, reaction time, memory, and feelings of fatigue.[24] One drawback is the predominance of research studies that only included young male participants.[37][1] Some meta-analyses find similar performance benefits in females to those found in males,[41][42][43] but more randomized controlled trials in females are needed to substantiate the generalization of sports nutrition recommendations for caffeine.[1][44]
Do different genotypes influence the effect of caffeine?
Does regular caffeine use reduce the benefit of caffeine on exercise performance?
Does regular caffeine use reduce the risk of cognitive disorders like dementia and Alzheimer's disease?
Is caffeine a banned substance for athletes?
Does coffee have the same effect as isolated caffeine on exercise performance?
Does using a caffeine-containing mouth rinse or chewing a caffeine gum have the same effect on exercise performance as ingesting caffeine?
Caffeine is generally safe for most people if used within the recommended amounts (up to 400 mg per day in a healthy adult, or up to 200 mg in a single dose — see the dosage information section.[3] Furthermore, caffeine-containing beverages like tea and coffee are socially acceptable and widely used.[45][12] However, caffeine has several side effects and drawbacks.
The side effects of caffeine include a short-lived rise in heart rate and blood pressure, heart palpitations, headache, increased urine output, nervousness, gastrointestinal problems, etc.[20][21][22][23] Caffeine can also raise heart rate during and following exercise.[46][2] Some of these side effects, particularly those related to heart rate, blood pressure, and urine output, may subside with regular use due to increased tolerance.[20][22][8][13][14] Because caffeine can cause mild drug dependence, some people also experience withdrawal symptoms — headache, drowsiness, and irritability — when they stop using caffeine after regular daily intake.[47]
One of the major drawbacks is that caffeine impairs sleep quality and can reduce sleep duration.[48][49] A recent meta-analysis found that to prevent deleterious effects on subsequent sleep, coffee should be consumed at least 9 hours before bedtime, and a caffeine-containing pre-workout supplement should be consumed at least 13 hours before bedtime.[48]
Sadly, there have been several case reports of caffeine toxicity[50][51][52][53] and caffeine-related death.[54][55][56][57] However, considering the high global prevalence of caffeine intake, these cases are rare.[45][12][58] Such cases also typically involve exceptionally high caffeine intakes (usually unintentional but sometimes purposeful) or caffeine intake combined with other drugs and alcohol.[54] Some instances of toxicity and death involve energy drinks — beverages that contain as much as 300 mg of caffeine plus other stimulants like taurine, in addition to B vitamins and sugar.[59] At the population level, energy drinks are unlikely to cause harm because there is a low prevalence of consumption.[60][61] However, in certain groups (children, teenagers, people with underlying heart conditions), health risks can emerge when people far exceed recommended daily caffeine intake limits by consuming energy drinks with other caffeine-containing foods and beverages.[60][61]
Some epidemiological studies have found a relationship between high daily caffeine intake (e.g. more than 4 to 6 cups of coffee per day) and increased cardiovascular disease risk.[62][63][64][65] However, the evidence from mendelian randomization studies and meta-analyses of epidemiological studies shows that caffeine (or coffee) intake does not increase the risk of cardiovascular disease, including coronary artery disease, stroke, heart failure, and cardiovascular disease mortality,[61][65][66][67] and that moderate intake might even be protective.[68][69][70][71]
Some studies also find a relationship between caffeine intake during pregnancy and poorer health outcomes for mother and baby following birth; however, the entirety of the evidence is equivocal.[72][73] Furthermore, it is unclear whether reducing caffeine intake during pregnancy has any effect on pregnancy outcomes, including infant health and birth weight.[72][73] However, some authorities recommend that pregnant women limit their caffeine intake to 200 milligrams per day (the equivalent of 1–2 cups of coffee per day or less).[5]
Caffeine’s mechanisms of action are not completely understood, but the proposed mechanisms likely act in unison.[14][1][74]
Caffeine’s primary action is in the central nervous system, where it prevents adenosine from binding to its receptor.[75][14][74] This affects the secretion of several neurotransmitters — norepinephrine, dopamine, acetylcholine, serotonin, GABA, etc. — involved in alertness, mood, motivation, memory, and pain perception.[14][76] For example, adenosine released during exercise decreases the secretion of dopamine, a neurotransmitter that increases mood and motivation.[1][77] So, when taken before or during exercise, caffeine allows dopamine secretion to continue, potentially maintaining mood and motivation to work hard.[77] This, coupled with lower pain perception caused by the lack of adenosine signaling, may partly explain caffeine’s sports-performance-enhancing effects.
Caffeine also plays a role in cellular calcium transport, which regulates neurotransmitter secretion in neurons and contractile force in muscle cells.[14][74] Caffeine can have a direct effect on calcium transport in muscle cells and may, therefore, have a direct effect on muscle contractile force,[1][78][79][80][81][82] providing an additional explanation for its performance-enhancing effects.
Caffeine can also inhibit a group of enzymes called cyclic nucleotide phosphodiesterases (PDEs), preventing the breakdown of cyclic adenosine monophosphate (cAMP).[14][74] This is important because cAMP stimulates the release of neurotransmitters like dopamine, epinephrine, and norepinephrine, which regulate aspects of cognitive function, including alertness, mood, motivation, and memory.[14][74] However, this mechanism requires more research and may only occur in the presence of high concentrations of caffeine, i.e., following high caffeine doses.
The optimal dose for sports performance benefits is 3–6 mg per kilogram of bodyweight (approximately 200–400 mg in a 70 kg person), taken around 60 minutes before exercise.[1] Using a dose at the low end of this range, approximately 3 mg/kg of caffeine before exercise (approximately 200 mg in a 70 kg person), achieves the benefits along with the lowest risk of side effects.[2]
Caffeine is generally safe for most people if used within the recommended amounts.[3][4] The US Food and Drug Administration (FDA) and the European Food Safety Authority (EFSA) report that a daily total caffeine intake of up to 400 milligrams (mg) does not raise safety concerns for non-pregnant adults.[5][6] The EFSA further reports that a daily total caffeine intake of up to 200 mg in pregnant and lactating women does not increase safety concerns for unborn babies or breastfed infants, and that a single caffeine dose of up to 200 mg (approximately 3 mg per kg bodyweight for a 70-kg adult) does not raise safety concerns.[5] In other words, caffeine intake is unlikely to cause detrimental health effects if daily intake is below 400 mg/day in non-pregnant adults, or below 200 mg/day in pregnant/lactating women, and if a single dose is less than 200 mg.
In children and adolescents, more research is needed to clarify safety, health risks, and tolerable upper intake limits.[5] Evidence from a systematic review suggests that children and adolescents should consume no more than 2.5 mg per kg of bodyweight per day (equivalent to 100 mg for a 40 kg person).[3] But some organizations discourage all consumption of caffeine and other stimulants by children and adolescents.[6]
Caffeine concentrations in various foods — coffee, tea, soft drinks (cola), chocolate, energy drinks, etc — are highly variable.[5] For example, a cup of coffee may contain approximately 50–200 mg of caffeine, but the amount per cup is highly variable depending on the type of bean, how it is roasted, and how it is prepared.[7][8] Some energy drinks contain very high amounts of caffeine, and the FDA recommends avoiding dietary supplements that contain pure or highly concentrated caffeine because a single teaspoon of pure powdered caffeine contains about 5 grams (5000 milligrams), far higher than the upper recommended limit of daily intake and approaching the toxic dose.[9]
For a detailed overview of caffeine dosing, read our article “How Much Caffeine Is Too Much?”.
Note that caffeine citrate is about 50% caffeine by weight, i.e., a 50 mg dose of caffeine citrate contains approximately 25 mg of caffeine.[10][11]
There are benefits associated with chronic caffeine consumption, and there are benefits associated with acute caffeine consumption that fade with tolerance; if you like the latter, cycling is mandatory. If you like the former, cycling is not needed.
Chronic non-cycled caffeine consumption (in the form of coffee and tea, both of which contain many bioactive components in addition to caffeine) is associated with a reduced risk of several diseases, including Parkinson’s disease, type 2 diabetes, and chronic liver disease.[61] Caffeine consumption has also been shown to enhance performance on attention tasks, regardless of the participants’ caffeine consumption habits.[114] Clearly, cycling isn’t needed to obtain these benefits. So why cycle? The question usually comes up in the context of exercise performance enhancement.
Supplementation with 3–6 mg of caffeine per kg of body weight before exercise has been consistently shown to enhance performance over a wide range of exercise intensities and durations, with beneficial effects on aerobic exercise performance, muscular endurance and strength, sprinting, jumping, and throwing performance.[1]
Caffeine elicits an ergogenic effect (i.e., enhances exercise performance) primarily by affecting the central nervous system. Caffeine blocks adenosine receptors in the brain, resulting in increased release of neurotransmitters such as dopamine and norepinephrine, and thus increased alertness and focus. It can also reduce pain and perceived exertion during exercise.
It seems to many habitual caffeine users that habitual use reduces, or even eliminates, the ergogenic effect of acute caffeine ingestion. The rationale is that habitual caffeine use increases the number of adenosine receptors in the brain, and as such, reduces the adenosine-blocking effect of caffeine. This has been shown in rodents,[115][116] but has yet to be studied in humans. Evidence from human studies does suggest that habitual caffeine use can blunt some aspects of the physiological response to caffeine, such as an increase in plasma epinephrine levels.[117][20]
In further support of the idea that athletes should cycle caffeine in order to maximize its ergogenic effect, some studies indicate that when individuals with low habitual caffeine intake (< 75 mg/day) consume 3 mg of caffeine per kg of body weight daily for 20–28 days, the ergogenic effect of caffeine is reduced.[93][94]
However, in a 2022 meta-analysis of 59 studies that investigated the effect of acute supplementation with caffeine on exercise performance in habitual caffeine consumers, it was found that acute supplementation with caffeine improved performance during endurance, power, and strength exercises.[97] Moreover, the performance-enhancing effects of an acute dose of caffeine were independent of whether or not that dose was greater or smaller than participants' habitual consumption, as well as being independent of sex or training status.
The results of this meta-analysis indicate that caffeine does not lose its ergogenicity with chronic use; that is, even with daily caffeine consumption, ingesting some caffeine about an hour before exercise will still have a positive impact on performance in people who benefit from caffeine (which is not everyone). But is it possible that skipping a cup of coffee today will enhance the effects of a cup of coffee on exercise performance tomorrow? Probably not.
Two separate studies reported that abstaining from caffeine for four days prior to an exercise test did not enhance the effect of acute supplementation with caffeine,[118][119] and shorter withdrawal periods (24–48 hours) don’t appear to be useful either.[97] Therefore, the available evidence does not support the common practice of utilizing a caffeine withdrawal period before an important training session or competition to maximize caffeine’s ergogenic effect. In fact, doing so may even be detrimental, as a four-day withdrawal period was found to result in a number of side effects, including headaches, fatigue, impaired focus, and a lack of motivation.
It remains to be determined whether a longer withdrawal period can boost caffeine’s ergogenic effect, but considering the fact that acute supplementation with caffeine enhances exercise performance in habitual users, and abstaining from caffeine tends to result in unfavorable symptoms, the juice is unlikely to be worth the squeeze if the purpose of a caffeine withdrawal period is solely to maximize caffeine’s ergogenic effect on a specific day.
Over the course of a day, you get sleepy as adenosine binds to A1 receptors in your brain. Caffeine blocks adenosine from binding, thus making you feel alert and also helping you feel better.
Caffeine is the most widely consumed psychoactive drug in the world, largely due to its mood-enhancing and stimulatory effects.
Despite widespread consumption, few people are actually aware of how caffeine works in the body.
We thought you might be interested in learning exactly how caffeine works in your body after you ingest it and it enters the brain.
The key player here is adenosine.
If you remember your high school biology, think of the lock-and-key model.
Adenosine is a key that opens up a variety of locks, with the locks actually being receptors in the brain.
Once adenosine (the key) locks into a certain receptor (the lock) in the brain, it has a unique effect on the brain.
There are a host of different receptors in your brain, so different ones have different effects. The one we’re interested in is the A1 receptor. Once adenosine locks with the A1 receptor, it promotes muscle relaxation and sleepiness, which is why people get tired as the day progresses.
Furthermore, adenosine can bind to the A2A receptor. When it binds, this interferes with the release of mood-improving neurotransmitters, such as dopamine.
Adenosine itself is produced primarily from physical work and intensive brain use. Thus, over the course of the day, your body accumulates adenosine.
If only there was something that could get in the way of adenosine from locking into the A1 receptor...
Adenosine is one of the best-known sleep-regulating molecules. Located in your central nervous system, it helps get you sleepy as the day winds down, among other effects.
When you first wake up, your body has metabolized away the adenosine molecules. You’re a bit groggy, but you’re waking up.
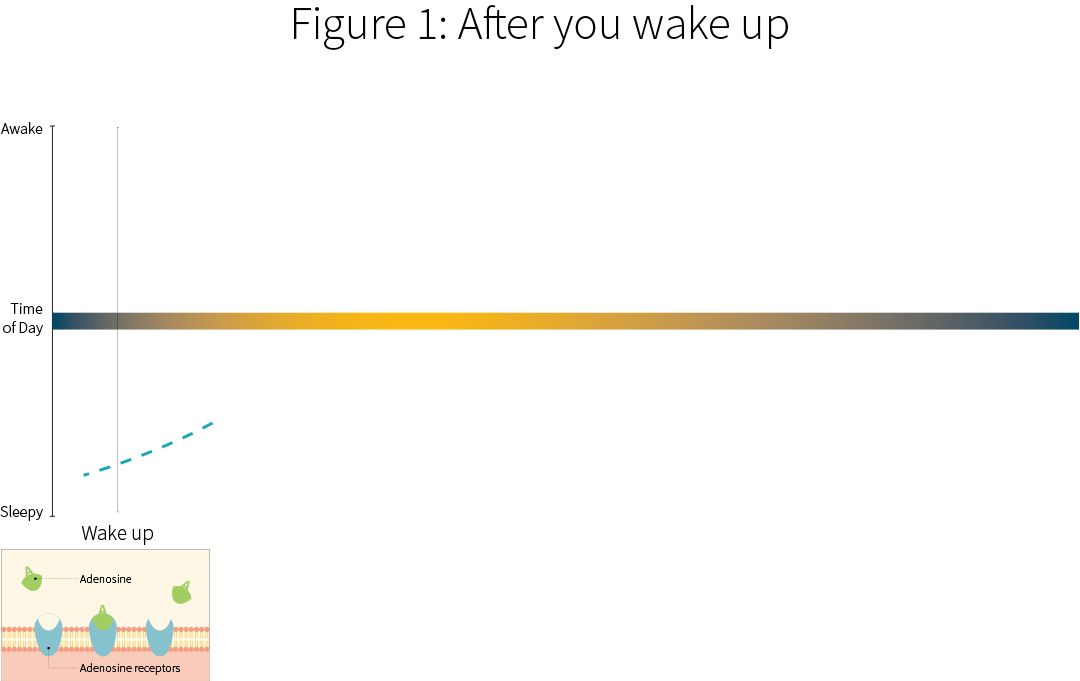
Most people initially drink caffeine in the form of a beverage. It’s absorbed in the small intestines within an hour, and becomes available throughout the blood and most parts of the body, including your brain.
As it starts entering your brain, it starts competing with adenosine.

Blood concentrations of caffeine tend to peak within two hours, which also means that brain concentrations of caffeine are at their peaks. The caffeine in your brain is competing with adenosine and preventing it from binding to A1 receptors. This is what gives you a jolt of wakefulness.
To be precise, the caffeine doesn’t actually lock in with the A1 receptor. It’s more like something that gets in the way and occupies the lock, rather than actually unlocking it.
It similarly gets in the way of the A2A receptor, which can help promote the release of dopamine and glutamate (and make you feel good after you drink coffee)!

Eventually, caffeine molecules will unbind from the adenosine receptors (as all molecules generally do).
Most of caffeine is metabolized through the CYP1A1/2 enzymes into various substances such as araxanthine, theobromine, and theophylline.
The half-life (the amount of time it takes for the concentration of a substance to be halved) of caffeine in the body ranges from three to ten hours depending on the amount of CYP1A1 in the body, which varies from individual to individual.

By early evening, most of the caffeine from your morning cup of coffee has metabolized. There are significantly fewer caffeine molecules occupying the A1 receptors, so adenosine starts binding to them.
This starts promoting muscle relaxation and sleepiness, and that’s why you start feeling sleepy.

When you go to sleep and your body starts recovering, the adenosine molecules are metabolized. This is why sleep is so important - one of the issues with a lack of sleep is the increase in adenosine molecules. This then takes us back to the "Before Caffeine" step.
Of course, you can always attempt to drink a larger dose of caffeine at one sitting, or drink caffeine multiple times during the day to keep sleepiness at bay. But that’s not really a sustainable strategy.
Caffeine allows people to remain awake by competing with a molecule that promotes sleepiness called adenosine. Caffeine has a similar shape to adenosine and prevents it from binding to its receptors.
For more information, check out our in-depth caffeine page.
As an alternative to ingesting caffeine, a small number of studies have tested the effects of chewing a caffeine-containing gum or mouth-rinsing a caffeine-containing solution. The evidence suggests that caffeine gum may have a very small beneficial effect on endurance and strength-related outcomes, but only if the gum is chewed less than 15 minutes before exercise at a caffeine dose of at least 3 mg per kilogram of bodyweight (equivalent to approximately 210 mg in a 70 kg person).[111] Meanwhile, the results from the small number of published mouth rinse studies are equivocal, so it is currently unclear whether mouth-rinsing caffeine is as effective as ingesting it.[112][113] Further high-quality research is needed to clarify whether these alternative routes of caffeine delivery are a valid alternative to ingesting caffeine.
Full page update
We updated this page and it's database with the newest meta-analyses.
One new meta analysis added
We added one additional meta-analysis to the database.
Reviewed By